Introduction
Cardiovascular diseases are known to be the leading cause of death worldwide. Many of them are associated with sclerotic, fibrotic, autoimmune and inflammatory processes in which the extracellular myocardial matrix plays the crucial role by determining the adaptive and regenerative capabilities of myocardium [1]. The myocardial extracellular matrix is not just a passive entity, but rather a complex and dynamic microenvironment which represents an important structural and signaling system within the myocardium [2, 3]. The network of collagen type I and III is associated with elastic fibers and binds the components of the myocardium together, retains the shape of the heart muscle and determines the mechanical properties of the myocardium during the cardiac cycle [4]. A number of authors have convincingly demonstrated that interstitial matrix plays the significant part in the progression of cardiac dysfunction, rupture and mortality [5, 6]. Morphologically such changes are represented by increased amount of collagen and fibrous tissue in the myocardium [5, 7]. Thus, ideas about the spatial organization of the intercellular substance of the myocardium remain controversial and are based mainly on stereological studies that do not allow to give a complete picture of the architectonics of the connective tissue component of the myocardium. Quantitative morphological studies of cellular elements of the myocardium require the search for various types of interstitial cells and their network.
The main cause of myocardial remodeling is chronic hypoxic state. One of the most widespread forms of oxygen deficiency is hypoxic or hypobaric hypoxia [8]. Industrial development of mountain regions, military, sports, recreational and other activities, development of the Arctic and mineral resources of Russian Federation pose challenges to applied biology and medicine related to the development of methods of selection, optimization of adaptation processes and preservation of high human performance in extreme environmental conditions, which is impossible without understanding the mechanisms of adaptation of the heart to intermittent hypobaric hypoxia [3]. In the cells exposed to hypoxia reactive oxygen species (ROS) damage membranes (including mitochondrial), exacerbate the disturbances of energy metabolism, i.e. a vicious circle is created [4, 7]. Understanding the fundamental role of hypoxia and peroxidation in the genesis of many cardiovascular diseases has stimulated the development of strategies that can enhance the energy-producing functions of cells [9]. In the treatment of the pathology of the cardiovascular system, a whole range of drugs with antianginal and antihypoxic properties are currently used but there is no reliable evidence obtained about their cellular and molecular effects [10, 11].
So, the aim of the current research is to study myocardial interstitial matrix with and without succinic acid treatment in the rats exposed to hypobaric hypoxia.
Material and Methods
Design of study
The study was conducted on 26 adult males of Wistar rats weighing 220-310 g, divided into 3 groups. Animals were housed in a vivarium and cared in accordance with the rules and regulations for the handling of laboratory animals. The first control group consisted of 6 intact animals. The second group included 10 rats which were exposed to hypobaric hypoxia without medication for 30 days. Third group was composed of 10 rats, which were medicated by succinic acid solution Cytoflavin which was injected intraperitoneally once a day at the rate of 0.5 ml/100 g of animal body weight 15 minutes before hypoxic exposition for 30 days [12]. A solution of 0.9% NaCl was administered intraperitoneally to the rats of the second group in a similar dosage.
Cytoflavin was provided by Scientific technological pharmaceutical company "POLYSAN" (Saint Petersburg, Russia). The active ingredients of Cytoflavin solution contain the following substances:
- Succinic acid 100 mg in 1 ml of solution,
- Inosine (Riboxin) 20 mg in 1 ml of solution,
- Nicotinamide (Vitamin PP) 10 mg in 1 ml of solution,
- Riboflavin mononucleotide (vitamin B2) 2 mg in 1 ml.
Rats were anesthetized with chloroform and sacrificed next day after last exposure. All efforts were made to minimize animal suffering and to reduce the number of animals used. Rats were sacrificed by decapitation under anesthesia (ether with chloroform) in accordance with the “International Recommendations (Code of Ethics) for conducting biomedical research using animals” (1985) and laboratory practice rules in the Russian Federation (order of the Ministry of Health of the Russian Federation from 19 June 2003, №267).
Animal experiment was approved by the Bioethics committee of Crimea Federal University Center (Protocol №8 from 15.03.2016) according to the permission of the Academic Council of the Crimean Medical Institute (No. 103 of 30.11.77). The research was approved by the Institutional Committee on Bioethics and is consistent with the International Guidelines for the Care and Use of Laboratory Animals published by the US NIH (No. 85-23, 1985) and Guide for the Care and Use of Laboratory Animals (2009).
Hypobaric hypoxia modeling
Throughout the experiment, animals of the second and third groups were daily immersed for 1 hour in a transparent hypobaric chamber equipped with a manometer, safety valve, alkaline absorber to eliminate excess carbon dioxide, where the pressure was below atmospheric, which corresponded to a rise of 6,000 m above sea level: (354.2 mmHg), which is equivalent to a moderately intense hypoxic exposure. The animals were housed in standard conditions with free access to food and water. Prolonged hypobaric hypoxia was modeled after determining the individual sensitivity, so we have used low-resistant rats.
Study of myocardial interstitial matrix
In the end of experiment the hearts of the above-mentioned males after thoraco-and pericardiotomy were removed and immediately placed in a cardioplegic solution (0.9% KCl at a temperature of 0°C) to achieve the relaxed myofibers due to the cardiac arrest in diastole. Fragments of right and left ventricle after sampling was immediately placed in 4% formaldehyde solution buffered to a pH of 7.2–7.4 with monosodium phosphate and processed through the usual technique for paraffin inclusion [13]. For the immunohistochemical study, sections were cut using the same equipment, but with a thickness of 3 μm. Sections were collected on poly-lysine coated slides, dried in a thermostat at 370 C for 24 hours in order to obtain a perfect adhesion of the biological material to the surface of the histological slide, and then stained using different antibodies: Anti-alpha smooth muscle Actin antibody (ab5694), Anti-MMP9 antibody [5G3] (ab119906). After antigen retrieval, sections were cooled down to room temperature and were incubated for 30 minutes in a 1% hydrogen peroxide solution. The sections were next washed in phosphate-buffered saline (PBS), followed by a blocking step of 30 minutes in 2% skim milk. Phosphate-buffered saline (abbreviated PBS) is a buffer solution commonly used in biological research. It is a water-based salt solution containing disodium hydrogen phosphate, sodium chloride and, in some formulations, potassium chloride and potassium dihydrogen phosphate. The osmolarity and ion concentrations of the solutions match those of the human body (isotonic). Next, the slides were incubated with the primary antibodies overnight at 400C, and the next day, the signal was amplified for 30 minutes using a peroxidase polymeric secondary detection system (EnVision, Dako).
Electron microscopy
Electron microscopy fixation of myocardium was performed by immersion in the mixture of 1.0% paraformaldehyde and 1.5% glutaraldehyde in 0.1 M phosphate buffer at PH 7.4. After fixation, the specimens were postfixed in a 1% osmium tetroxide solution, and then washed 3 times in the buffer solution, continuing with dehydration from 50% ethanol, then with uranyl acetate (2% in 60% ethanol) with ascending graded ethanols up to 100%, and then washed in propylene oxide [14]. They were embedded in an Epon-Araldite composite, after those semithin sections and ultrathin sections were cut by the ultra-microtome UltraCut. Semithin sections were stained with tolouidine blue and ultrathin sections, with lead citrate, and examined on SELMI electron microscope.
Statistical analysis
Differences in response of the experimental groups to hypoxia were revealed by the quantitative analysis of the extracellular (stromal) components of the myocardium. We performed 30 measurements for every morphometrical index, that made 90 measurments for every animal group. Statistical data were analyzed using MS Excel (Microsoft, USA) and Statistica 10.0 (Statsoft, USA). Data distribution was normal according to the results of Kolmogorov-Smirnov test. Thus, data presented as mean with standard deviation and standard error of mean – M±SD (±SEM). Parameters between groups were compared by Student's t-test. p<0.05 were considered as statistically significant.
Results
Interstitial matrix and capillary measurements
The routine microscopic studies have revealed that in the second group of rats with no treatment the quality analysis of microslides showed that in contrast to the control group almost the entire stroma was represented by the thickened collagen fibers and proliferating cells of the connective tissue. Their total number increased relatively to the control by 31% initiating the fibrotic processes which may lead to cardiosclerosis if hypoxic exposure continues. Hypoxic effects in the third group receiving treatment were not significantly different from control according to the observed changes in the stroma. Most of it demonstrated the dilated capillaries and edematous intercellular space. The number of connective tissue cells was not significantly different from control (Table 1).
Table 1. Morphometric analysis of myocardial interstitial matrix in rats expose to hypobaric hypoxia
|
Parameters |
Control group (n=6) |
Hh group (n=10) |
Hh + Cytoflavin group (n=10) |
|
Number of nucleated cells in the interstitial matrix |
31.20±0.87 (±0.36) |
41.90±0.91 (±0.37)* |
32.70±1.42 (±0.57) |
|
Average area of stromal cells nuclei, μm2 |
12.37±4.25 (±1.72) |
11.13±4.38 (±1.79) |
9.37±3.54 (±1.45)* |
|
Average area of interstitial matrix, μm2 |
328.35±40.49 (±16.53) |
262.90±35.46 (±14.48)* |
341.13±45.91 (±18.76) |
Data presented as mean with standard deviation and standard error of mean – M±SD (±SEM).
* – statistically significant (p<0.05) difference with the similar parameters in the control group. Hh, hypobaric hypoxia.
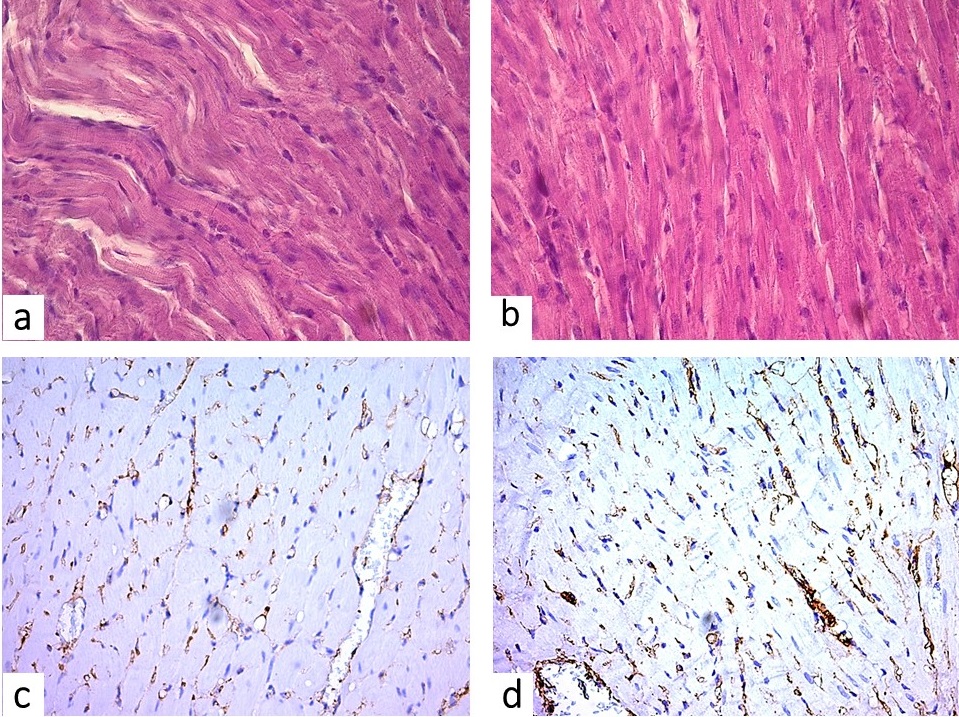
Figure 1. Rat myocardium general overview: (a) – swelling of contractile cardiomyocytes, signs of cell dystrophy, interstitial edema (H&E, x400); (b) and (c) – CD31 expression showing reduction of capillary patters (x400); (d) – CD31 expression demonstrates abundant capillaries (x400).
Other structural changes of the myocardium were represented by swelling of contractile cardiomyocytes, homogenization of their sarcoplasm (Figure 1a). In polarization microscope cells had fuzzy contours, they did not trace transverse striation, there was a marked disintegration of myofibrils, overrelaxation of sarcomeres, and some areas demonstrated myocytolysis. The capillary network was heterogenous: areas of constriction alternated with the dilatations of the vessel’ lumens with blood plasma diffusing through their walls followed by perivascular edema and erythrocyte diapedesis into the perivascular space (Figure 1с). Separate groups of cardiomyocytes were thinned, their sarcoplasma was unevenly stained. There were also signs of intermuscular edema.
The administration of succinic acid combined with inosine and vitamins diminished the dystrophic and necrobiotic changes of cardiomyocytes in the third group of rats. Histological examination of myocardial samples showed diminished groups of some muscle fibers, homogenization of sarcoplasma and reduction of glycogen granules. Vacuolization of cardiomyocyte sarcoplasm occurred in few cases. Most of the cardiomyocytes had vivid cell boundaries, uniformly stained sarcoplasm, large nuclei (Figure 1b). Polarizing microscopy traced clear myofibrils, intercalated discs. Though in cardiomyocytes with signs of destruction, I-II degree contractures were detected, less frequently there was a marked collapse of the myofibrils.
Meanwhile, after Cytoflavin administration it was found that in the third group of the walls of some vessels were thickened, endothelial cells protruded into the lumen in the form of a paling. In some vision fields a discontinuity of the vascular pattern was observed (Figure 1d). The capillary diameter exceeded the measurment of the second group (p<0.05), which indicates the relative safety of the hemomicrocirculatory vessels but was significantly lower than the control value (p<0.05). Accordingly, to the series without correction, the cross-sectional area of the capillaries was increased up to 28.44±0.34 μm2 against 24.53±0.49 μm2 (p<0.05). The total cross-sectional area of the capillaries is 0.77±0.24 x103 μm2. The relative surface area of the vascular area was lower than the control one by 21.8% (p<0.05) (Table 2).
Table 2. Microcapillary measurements in rats exposed to hypobaric hypoxia
|
Parameters |
Control group (n=6) |
Hh group (n=10) |
Hh + Cytoflavin group (n=10) |
|
Diameter of the capillary, μm |
8.52±0.29 (±0.12) |
5.59±0.17 (±0.07)* |
6.02±0.18 (±0.07)* |
|
The cross-sectional area of the capillary, μm2 |
56.98±0.56 (±0.23) |
24.53±0.49 (±0.20)* |
28.44±0.34 (±0.14)* |
|
The numerical density of the capillaries, N |
30.32±0.39 (±0.16) |
27.51±0.83 (±0.34)* |
27.15±0.39 (±0.16)* |
|
Total cross-sectional area of cross sections of capillaries, x103 μm2 |
1.73±0.47 (±0.18) |
0.67±0.26 (±0.10)* |
0.77±0.24 (±0.10)* |
|
Relative superficial area of vessels, % |
31.60±0.29 (±0.12) |
21.50±0.44 (±0.18)* |
24.70±0.56 (±0.23)* |
Data presented as mean with standard deviation and standard error of mean – M±SD (±SEM).
* – statistically significant (p<0.05) difference with the similar parameters in the control group. Hh, hypobaric hypoxia.
Fibrogenesis in the interstitial space
After the exposure to hypobaric hypoxia lysis of single cardiomyocytes was noted in second group rat myocardium. A proliferation of connective tissue fibers was noted at the site of the dead cardiomyocytes (Figure 2a) along the capillaries and around the larger vessels (Figure 2c). This was due to the fact that in response to ischemia fibroblast activation occurs resulting in an increase in collagen synthesis. A disproportionate increase in the synthesis and inhibition of the collagen degradation in the extracellular matrix can cause subsequent fibrogenesis — a disproportionate accumulation of fibrillar collagen.
Third experimental group cardiac interstitial matrix demonstrated mild edema around the arterioles and venules. Electron microscopy revealed many fibroblasts were involved in myocardial tissue compartmentalization, separating certain groups of cardiomyocytes from adjacent capillaries without overproduction of collagen fibers (Figure 2b). Many fibroblasts had formed thin plate-like processes separating cardiomyocytes from adjacent capillaries, as well as adjacent bundles of cardiomyocytes from each other. However, multiple foci of newly formed connective tissue were also detected near necrotic cardiomyocytes and around capillaries (Figure 2d). Hypertrophied cardiomyocytes with large nuclei were located nearby to maintain the pumping blood volume and compensate the previous loss of cardiac cells by launching the internal compensatory program aimed at adapting functioning cardiomyocytes to increased workload.
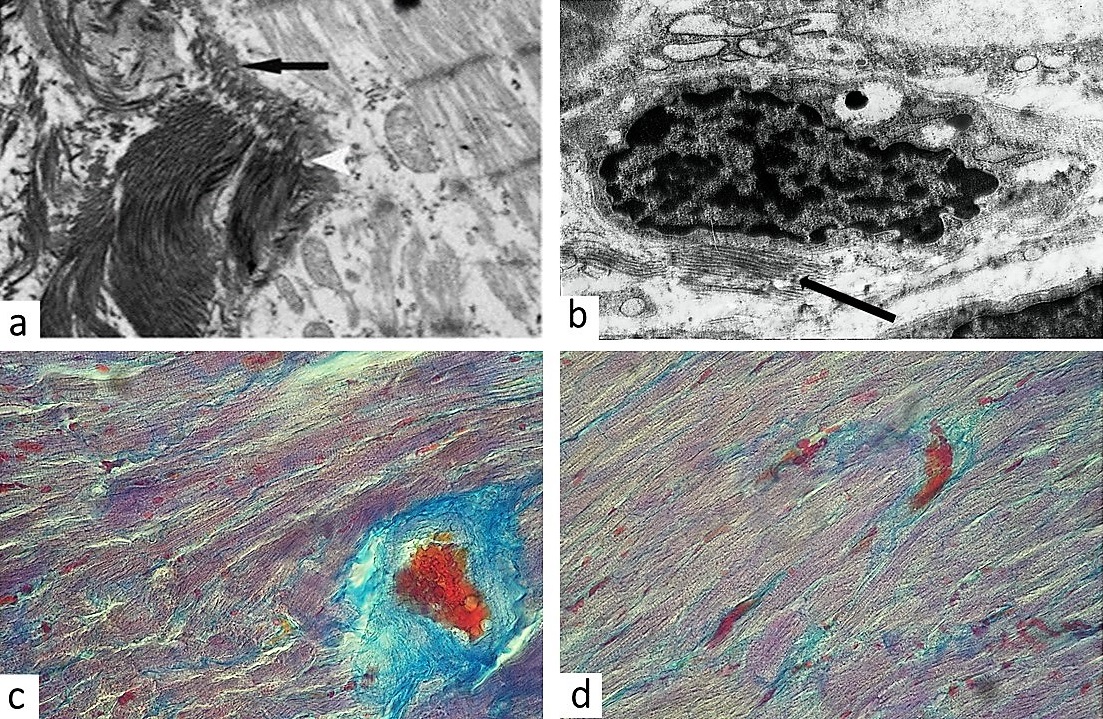
Figure 2. Collagen deposition in the cardiac interstitium: (a) – collagen synthesis replaces lesioned cardiomyocyte (arrows) (TEM, x 12,000); (b) – fibroblast with low amount of collagen fibers (arrows) interconnected with sarcolemma (TEM, x12,000); (c) – blue connective tissue increases the distance required for diffusion of oxygen between cardiomyocytes and arteriole (Mallory staining, x400); (d) – blue connective tissue between cardiomyocytes and capillaries (Mallory staining, x400).
Markers of the initial myocardial remodeling
An increase in the number of myofibroblasts which was observed in the hypoxic rat heart in the second group can affect the ability of fibroblasts to conduct polarization from one cardiomyocyte to another (Figure 3a). However, myofibroblasts can maintain dynamical features of myocardium by maintaining myocardial integrity against a background of continuous mechanical forces associated with the heart pumping. This effect was probably observed in the third group because fibroblasts were located mainly nearby hypertrophied cardiomyocytes integrating their contractions with another myocardial element (Figure 3b). Still even moderate activation of fibroblasts can lead to progressive fibrogenesis.
Prolonged hypobaric exposure triggered activation of intensely colored connective tissue cells (neutrophils, macrophages and fibroblasts) located in perivascular and interstitial space. These cells were expressing the matrix metalloproteinase-9 (MPP-9), the intensity of IHC staining was high, i.e. there was a complete uniform staining of the cytoplasm (Figure 3c). It is known that the sustained high concentrations of MMP-9 testify to the permanence of the destructive processes in the extracellular matrix of the myocardium and are predictors of poor prognostic outcome of the disease.
In rats of the third group only single MMP-9-positive cells were detected in the specimens at separate sites in the myocardium, the severity of expression of the marker was slightly positive indicating the low intensity of the myocardial remodeling processes (Figure 3d).
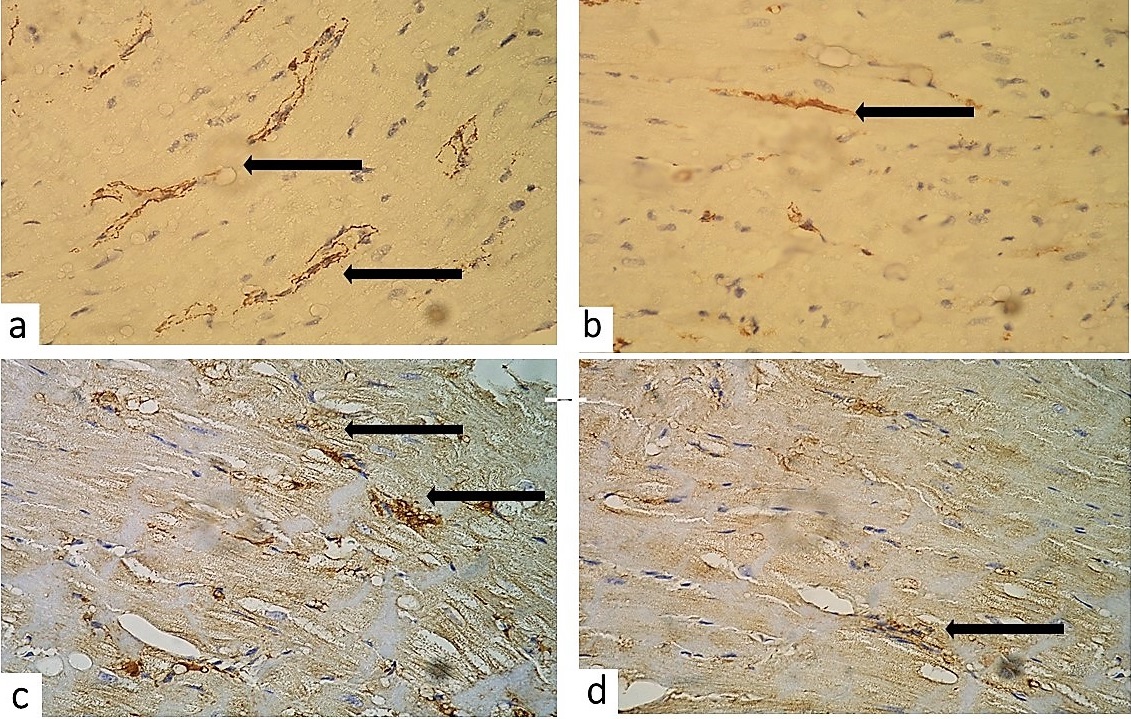
Figure 3. Identification of the main trigger proteins of myocardial remodeling in rats exposed to hypobaric hypoxia: (a) – the presence myofibroblasts in the expanded perivascular and interstitial spaces (x400); (b) – few myofibroblasts adjacent to enlarged cardiomyocytes (x400); (c) – myocardium of male rats exposed to histotoxic hypoxia (high cytoplasmic expression of MPP-9 by interstitial cells, x400); (d) – myocardium of male rats exposed to hypobaric hypoxia with subsequent correction (slightly positive expression of MPP-9 by interstitial cells, x400).
Discussion
The connective tissue extracellular matrix contains a network of macromolecular fibers supporting cardiomyocytes. It has a complex spatial organization that largely determines the structural and functional integrity of the heart muscle. In quantitative ratio the cellular elements of the connective tissue (65-75%) predominate in the myocardium, ensuring that the cardiomyocytes perform their main contractile function [15]. Myocardial fibrosis can be defined by a significant increase in the collagen volume fraction of myocardial tissue, is always present in end-stage heart failure. The distribution of myocardial fibrosis, however, varies according to the underlying pathology and accounts for discrepancies among different pathological reports in which only qualitative as opposed to quantitative measurements were made. Different types of myocardial fibrosis have been reported according to the cardiomyopathic process Reparative (compensatory) fibrogenesis develops at the site of the dead cardiomyocytes [16]. In return the reactive fibrogenesis we observed in our investigation was not directly related to cardiomyocyte necrosis, was presented as interstitial (accumulation of collagen in the intermyofibrillar space) and perivascular (accumulation of collagen within the advent of coronary arteries and arterioles) fibrogenesis [11].
Undoubtedly developing fibrogenesis has a negative impact on the state of the myocardium, contributing to the remodeling process. The accumulation of collagen causes an increase in the rigidity of the ventricular wall, which leads to a violation of contractility and relaxation [7]. The progressive accumulation of connective tissue leads to a decrease in capillary density and increases the distance required for diffusion of oxygen, which can determine the development of subsequent ischemia of cardiomyocytes. Fibrosis can also disrupt the electrical interaction between cardiomyocytes contributing to the development of arrhythmias [1].
The vast majority of cells of the connective tissue component of the myocardium are fibroblasts (90-95%). They are the main producers of matrix macromolecules, including collagen and major structural proteins [18]. The remaining cell forms are mast cells, macrophages, lymphoid cells. Thus, among fibroblasts, activated fibroblasts (collagenoblasts), fibroblasts, and myofibroblasts are isolated.
Myofibroblasts combine the qualities of a fibroblast (collagen synthesis, mainly of the III type) and a smooth myocyte (the presence of myofilament, the ability to contract). Myofibroblasts are attributed to the ability of dynamic changes in the volume of the intercellular substance, as well as the effect on the orientation of its fibrous elements [16] The switching of fibroblast phenotype to myofibroblasts is due to a transforming growth factor beta-1 (TGF-pi) and is associated with the beginning of their expression of smooth muscle alpha actinin (a-SMA) and desmin [5]. Appreciating the dual roles of cardiac myofibroblasts in the myocardial remodelling process is important, as they can be perceived to be both beneficial and detrimental according to the amount and their temporal and spatial location.
Fibroblasts secrete the proteins of the myocardial extracellular matrix and matrix metalloproteinases, thereby performing central role in maintaining the structure of the stroma. Collagen degrading enzymes belong to the group of matrix metalloproteinases that can cleave a large number of proteins of the extracellular matrix [19]. Matrix metalloproteinases have some similar properties: they share common regions of the amino acid sequence, are synthesized as inactive pro-enzymes, and require zinc as a cofactor [20]. The balance between the synthesis of matrix proteins and their degradation plays an essential role in maintaining the integrity of the myocardium. They are secreted in an inactive form into the extracellular space, where they are activated by the other proteases and participate in tissue remodelling. It is known that the progressive activation of matrix metalloproteinases in ischemia promotes dilatation and deterioration of left ventricular function that leads to the development of heart failure [21].
Cyto- and angioprotective effects in rats after medication with succinic acid combined with inosine, riboflavin and nicotinamide in complex therapy are associated, in our opinion, with the drug composition where active components have mutually potentiating effects and work as inducers of the main metabolic pathways in cells, and activators of key energy-forming processes. Inosin (Riboxin) is the agonist of purinergic receptors that activate the glycolysis. Inosin-induced activation of glycolysis helps to maintain the energy balance in the cell [10, 11]. Riboflavin (vitamin B2) is a coenzyme of flavin adenine dinucleotide (FAD) that activates succinate dehydrogenase and other redox reactions of the Krebs cycle. Nicotinamide (vitamin B3) in cells, through a cascade of biochemical reactions, is transformed into the form of nicotinamide adenine nucleotide (NAD) and its phosphate (NADP), activating the nicotinamide-dependent Krebs cycle enzymes necessary for cellular respiration and stimulation of ATP synthesis.
Conclusion
The analysis of the current reviews allows us to state that activated fibrosis is an essential component of myocardial remodeling during hypobaric hypoxia. Fibrosis in the myocardium inevitably leads to increased myocardial stiffness, resulting in systolic and diastolic dysfunction, neurohormonal activation and, ultimately, heart failure. Revealing the alterations in cardiac metabolism and function associated with sustained exposure to high altitude advances our understanding of hypoxia-related disease.
Succinic acid combined with inosine acts as a high-energy phosphate reserve, to maintain ATP at levels sufficient to support contractile function. These changes are probably associated with the positive effect of cytoflavin on the energy processes in the cell with the prevention of excessive production of reactive oxygen species, which improves the state of the endothelium and, therefore, the hemodynamics of the myocardium. It is obvious that the components of the drug as an integral part of a particular physiological system can affect the molecular targets of myocardial remodeling, apparently due to the activation of endogenous inhibitory factors that can suppress the processes of excessive fibrogenesis.
Limitations
The present experimental study was performed on young Wistar rats exposed to multiple episodes of hypobaric hypoxia over a period of 30 days. In order to that fact the study is limited only to those experimental conditions that have been modeled. There are reasonable limitations related to the extrapolation of the obtained data to the humans taking into account the species differences in the stress reaction and adaptive reserves.
Conflict of interest
The authors declare that they have no conflict of interest.
Ethical Approval
All applicable international, national, and institutional guidelines for the care and use of animals were followed. All procedures performed in the study involving animals were in accordance with the ethical standards of the institution or practice at which the study was conducted. This article does not contain any studies with human participants performed by any of the authors.
- Gammella E, Recalcati S, Rybinska I, Buratti P, Cairo G. Iron-induced damage in cardiomyopathy: oxidative-dependent and independent mechanisms. Oxid Med Cell Longev 2015; 2015: 230182. https://doi.org/10.1155/2015/230182.
- Burlew BS, Weber KT. Connective tissue and the heart. Functional significance and regulatory mechanisms. Cardiol Clin 2000; 18(3): 435-442. https://www.ncbi.nlm.nih.gov/pubmed/10986582.
- González A, Schelbert EB, Díez J, Butler J. Myocardial interstitial fibrosis in heart failure. J Am Coll Cardiol 2018; 71(15): 1696-1706. https://doi.org/10.1016/j.jacc.2018.02.021.
- Murphy-Ullrich JE, Sage EH. Revisiting the matricellular concept. Matrix Biol 2014; 37: 1-14. https://doi.org/10.1016/j.matbio.2014.07.005.
- McCurdy S, Baicu CF, Heymans S, Bradshaw AD. Cardiac extracellular matrix remodeling: fibrillar collagens and Secreted Protein Acidic and Rich in Cysteine (SPARC). J Mol Cell Cardiol 2010; 48(3): 544-549. https://doi.org/10.1016/j.yjmcc.2009.06.018.
- Frangogiannis NG. Matricellular proteins in cardiac adaptation and disease. Physiol Rev 2012; 92(2): 635-688. https://doi.org/10.1152/physrev.00008.2011.
- Cerit L. SYNTAX score and left ventricular remodelling. Int J Cardiol 2016; 223: 465. https://doi.org/10.1016/j.ijcard.2016.08.180.
- Bartsch P, Gibbs JS. Effect of altitude on the heart and the lungs. Circulation 2007; 116(19): 2191-2202. https://doi.org/10.1161/CIRCULATIONAHA.106.650796.
- Boylston JA, Sun J, Chen Y, Gucek M, Sack MN, Murphy E. Characterization of the cardiac succinylome and its role in ischemia-reperfusion injury. J Mol Cell Cardiol 2015; 88: 73-81. https://doi.org/10.1016/j.yjmcc.2015.09.005.
- Ariza AC, Deen PM, Robben JH. The succinate receptor as a novel therapeutic target for oxidative and metabolic stress-related conditions. Front Endocrinol (Lausanne) 2012; 3: 22. https://doi.org/10.3389/fendo.2012.00022.
- Hoffmeister HM, Betz R, Fiechtner H, Seipel L. Myocardial and circulatory effects of inosine. Cardiovasc Res 1987; 21(1): 65-71. https://www.ncbi.nlm.nih.gov/pubmed/3664538.
- Zadnipryanyi IV, Tretyakova OS, Sataeva TP. Investigation of the antioxidant activity and cardioprotective effect of reamberin and cytoflavin in newborn rats exposed to chronic hemic hypoxia. Arkh Patol 2015; 77(6): 39-44. Russian. https://doi.org/10.17116/patol201577639-44.
- Merkulov GA. Course of pathological/histological technique. Leningrad: Meditsina, 1969. 423 p. Russian. https://search.rsl.ru/ru/record/01007362447.
- White K. Electron microscopy: methods and protocols (3rd Ed.). Kuo John (Ed.). Humana Press, Totowa, NJ, 2014,799 pages. ISBN: 9781627037754. Microsc Microanal 2014; 20(5): 1624. https://doi.org/10.1017/s143192761401321x.
- Schwartzkopff B, Mundhenke M, Strauer BE. Remodelling of intramyocardial arterioles and extracellular matrix in patients with arterial hypertension and impaired coronary reserve. Eur Heart J 1995; 16 Suppl I: 82-86. https://www.ncbi.nlm.nih.gov/pubmed/8829962/.
- Pinto AR, Ilinykh A, Ivey MJ, Kuwabara JT, D'Antoni ML, Debuque R, et al. Revisiting cardiac cellular composition. Circ Res 2016; 118(3): 400-409. https://doi.org/10.1161/CIRCRESAHA.115.307778.
- Fan D, Takawale A, Lee J and Kassiri Z. Cardiac fibroblasts, fibrosis and extracellular matrix remodeling in heart disease. Fibrogenesis Tissue Repair 2012; 5(1): 15. https://doi.org/10.1186/1755-1536-5-15.
- Chen B, Frangogiannis NG. Macrophages in the remodeling failing heart. Circ Res 2016; 119(7): 776-778. https://doi.org/10.1161/CIRCRESAHA.116.309624.
- Kiczak L, Tomaszek A, Bania J, Paslawska U, Zacharski M, Janiszewski A, et al. Matrix metalloproteinase 9/neutrophil gelatinase associated lipocalin/tissue inhibitor of metalloproteinases type 1 complexes are localized within cardiomyocytes and serve as a reservoir of active metalloproteinase in porcine female myocardium. J Physiol Pharmacol 2014; 65(3): 365-375. https://www.ncbi.nlm.nih.gov/pubmed/24930508.
- Herpel E, Singer S, Flechtenmacher C, Pritsch M, Sack FU, Hagl S, et al. Extracellular matrix proteins and matrix metalloproteinases differ between various right and left ventricular sites in end-stage cardiomyopathies. Virchows Arch 2005; 446(4): 369-378. https://doi.org/10.1007/s00428-004-1177-z.
- Wells JM, Gaggar A, Blalock JE. MMP generated matrikines. Matrix Biol 2015; 44–46: 122-129. https://doi.org/10.1016/j.matbio.2015.01.016.
Received 28 November 2018, Revised 30 March 2019, Accepted 09 April 2019
© 2018, Zadnipryany I.V., Sataieva T.P., Tretiakova O.S., Zukow W.
© 2018, Russian Open Medical Journal
Correspondence to Igor V. Zadnipryany. Address: 5/7, Lenin Avenue, Simferopol, 295051, Russia. E-mail: zadnipryany@gmail.com.