Introduction
Hypoxia, on the one hand, can lead to the development of inflammation, but on the other hand, any inflammatory process especially with pronounced system manifestations is accompanied by oxygen deficiency [1-2]. It is known, that hypoxia plays a key role in both the system and local inflammatory reactions, mainly due to microcirculatory disorders and disseminated intravascular coagulation [3]. In this regard a number of studies have shown that severity of any inflammatory diseases, including inflammatory diseases of the gut such as Crohn's disease (CD) and ulcerative colitis (UC) depends on hypoxia resistance [4-7].
UC, CD and other intestinal diseases of unknown etiology on the clinical manifestation, mechanisms of their development and treatment approaches are combined into the group of inflammatory bowel diseases (IBD) [4,5,8]. In CD, all parts of the gastrointestinal tract may be affected, while in UC, pathological process develops first in the rectum and then spread proximally, affecting all part of the colon [9].
IBD is widespread throughout the world, and UC happens more often than CD. According to the statistic, the highest rate of UC cases is in North Europe and North America, where they range from 156 to 291 on 100,000 people [10]. The lowest rate is determined in African and Asian countries. In Russia frequency of UC is 20 cases per 100,000 people, CD is 3.5 per 100,000 people. In IBD the risk of colorectal cancer increases and its frequency is 0.4-0.8% [4].
The factors contributing the IBD development include environmental influence, tobacco smoke, oral contraceptives, titanium dioxide in toothpastes, microplastics particles etc. [4-5]. However accurate evidences about relation of these factors with mortality have not been revealed. It is currently believed, that the onset of IBDs associates with genetic predisposition [5]. The key loci associated with UC and CD development were identified; this is nucleotide-binding oligomerization domain containing 2 (NOD2), which is an intracellular receptor of immune cells involved in innate immunity reactions [11], macrophage-stimulating protein-1 (MST1), that regulates expression of Foxp3 and, therefore, the expression and development of regulatory T-lymphocytes (Tregs) [12], and finally the major histocompatibility complex (MHC), which is central for the presentation of peptide antigens to T-cells [13]. Mutations in loci of NOD2, 3p21 (MST1) and MHC are also detected in patients with such immune related diseases as ankylosing spondylitis and psoriasis [8].
During the last few years, important roles in the development of IBD play the changes in microflora composition. It was demonstrated that Clostridium difficile lead to a fulminant course of pseudomembranous colitis in humans, because they produce toxins, such as A-enterotoxin, which disturbed the barrier function of the intestinal mucosa, and B-cytotoxin [14]. Due to the increase in mucosal permeability, obligate and conditionally pathogenic microflora translocates to the intestinal wall, lymph nodes and other organs, which results in the increase of lipopolysaccharides level, which are the classic pro-inflammatory endotoxins and, which through TLR4 and NF-kB increase the production of pro-inflammatory cytokines, so that the cellular and humoral immune responses are activated [15].
Thus, IBD is a group of difficult to diagnose widespread among the population disorders. Now there are about 1-1.4 million people with IBD in the USA. Pathogenesis of the disease is associated with a complex interaction of the genetic factors, the environment, the microbiome and the uncontrolled reaction of the immune system, and the existing treatment methods are not effective enough.
Hypoxia-inducible factor
During hypoxia, to prevent the lack of oxygen in the organism, a complex repertoire of transcriptional changes is being realized, which, on the one hand, is targeted to the reduction of oxygen consumption, and on the other hand, to enhancement its delivery to cells or tissue. Hypoxia-inducible transcription factor (HIF) has a central role in the regulation of this transcriptional response to a decrease in oxygen content. HIF consists of an α-subunit sensitive to oxygen (HIF-1α, HIF-2α, HIF-3α) and a constitutive co-activator HIF-1β. HIF-1α and HIF-2α can regulate both common genes and also have pro- and anti-inflammatory effects on the expression of different genes [16]. HIF is responsible for the activation of more than 200 genes involved in erythropoiesis, angiogenesis, intestinal barrier integrity, iron homeostasis and glycolysis [17].
Under normoxic conditions, mRNA of HIF-α is expressed constitutively, but the HIF protein is rapidly and efficiently disintegrates due to the activity of the prolyl hydroxylase (PHD) related to the 2-oxoglutarate-dependent dioxygenase family. If there is a sufficient level of oxygen, PHDs hydroxylate specific proline residues in the oxygen-dependent domain of HIF-α [18-19], directing the protein to VHL-dependent (tumor suppressor Von Hippel– Linda) ubiquitination and proteasome degradation [20].
During hypoxia, the ability of PHD to hydroxylate HIF-α is impaired, since atomic oxygen is used in the reaction. Then HIF-α can accumulate and translocate into the nucleus, where it activates gene expression as a result of stabilization with HIF-1β and p300.
Microcirculation of blood in the intestines
Oxygenated blood enters the intestine from the celiac trunk, the upper and lower mesenteric arteries which makes up 20-25% of cardiac output in fasting conditions, but increases dramatically in response to food intake. Nutrients such as glucose, peptides and lipids can increase total intestinal blood flow by more than 200%. However the partial pressure of oxygen (pO2) in the perivascular zones at the apex of the villus can be reduced by about half under the same conditions [21]. The intestinal mucosa is well vascularized, however, the villi of the small intestine and the crypt of the large intestine receive a disproportionate amount of blood flow — 60% in the villi of the small intestine, 40% in the colon crypts [22].
The villi of the small intestine are perfused by the bringing and outflowing vessels, which form a capillary loop at the top of the villi, before blood enters the venule. Considering the size of the villus (<20 µm in diameter) and the combination of arterial and venous blood flow in this small physical space, there is a possibility for a backflow of oxygen shunt between vessels running in opposite directions. This includes the oxygen diffusion in the villus, directly into the venule, thus bypassing the normal vascular contours of the villi [23]. The consequence of this is a high pO2 gradient in the villi in the zone of the crypts and the apex of the villi. The blood supply decrement of the mucosa may contribute to a more pronounced decrease in pO2 on the surface of the intestine.
Physiological hypoxia of the intestine
The epithelial barrier is located on the border between the internal and external environment of the organism and consists of the mucus layer, the glycocalyx and the epithelial lining [24]. The epithelial lining is formed by a single layer of cylindrical cells represented by colonocytes or columnar absorptive cells, the apical part of which forms the lining of the luminal surface of the intestine, by the goblet cells, enteroendocrine and M-cells [25]. Intestinal epithelial cells (IECs) are interconnected by a complex of intercellular contacts that maintain the integrity of the epithelial lining and prevent the paracellular transport of bacteria and macromolecules [24]. IECs are covered with mucus, which provides protection from physical and chemical damage.
IECs are supplied with oxygen from the vessels of the microvasculature, depending on their localization on the surface of the villus or crypt. In addition, the oxygenation of ESCs is depended on their location relative to the oxygen environment of the intestinal lumen.
The role of the intestinal microbiota in the conditions of the physiological norm and in the mechanisms of the development of a wide range of diseases, such as IBDs, autism, obesity, Alzheimer's disease and others, is being actively studied [25-28]. The main functions of the intestinal microbiota include competition with pathogenic bacteria (for example, Clostridium difficile), synthesis of vitamins (for example, vitamin K), modification of bile acids, and the production of short-chain fatty acids (SCFA), such as butyrate, propionate and acetate [26]. According to the literature, butyrate increases the microbiota's oxygen consumption and HIF stabilization in mouse IECs, while HIF expression in IECs of gnotobiotic mice is reduced and hypoxia-sensitive dyes in these mice are delayed [29]. All of these indicates the important role of microbiota in the establishment of "physiological" hypoxia, depending on the content of butyrate. F. Rivera-Chávez et al. [30] observed an increase in colonocyte oxygenation in response to depletion of producing butyrate Clostridia pool. Surprisingly the elimination of commensal anaerobic Clostridia with antibiotics contributes to the enhanced growth of Salmonella in the intestine [30].
Thus, the intestinal mucosa appears to be in conditions of "physiological hypoxia". The degree of intestinal oxygenation is influenced by the oxygen diffusion from subepithelially located capillaries, the localization of IECs in the villi and crypts, the change in the anaerobes and aerobes ratio, and the production of SCFA (for example, butyrate) by microorganisms.
HIF and ulcerative colitis
There is a disturbance of the tight junctions integrity in IBDs, that increases the permeability of the epithelial barrier, and a destruction of the mucins, forming the biofilm in which the commensal microflora is located, leads to the pronounced antigenic stimulation of the intestinal mucosa immune cells. HIF-1α directly regulates the activation of several protective genes in response to the damage to the epithelial barrier integrity, reduces the production of cytokines and leads to the increase in production of β-defensins (Figure 1), that play a key rolein antimicrobial immunity [31]. HIF-2α directly regulates the production of some pro-inflammatory cytokines, including tumor necrosis factor-α, which leads to HIF-2α-induced inflammation [32]. In addition, a prolonged increase in the level of HIF-2α results in a high turnover of tight junction proteins, such as occludin, which leads to a decrease in the integrity of the epithelial barrier [33] (Figure 1). It was shown that HIF-2α plays an important role in reparative processes in wound inflammation [32-33]. It is important to note that overexpression of both HIF-1α and HIF-2α genes enhances the inflammatory response, which suggests that activation of HIF-1α does not prevent the pro-inflammatory response, induced by HIF-2α [32].
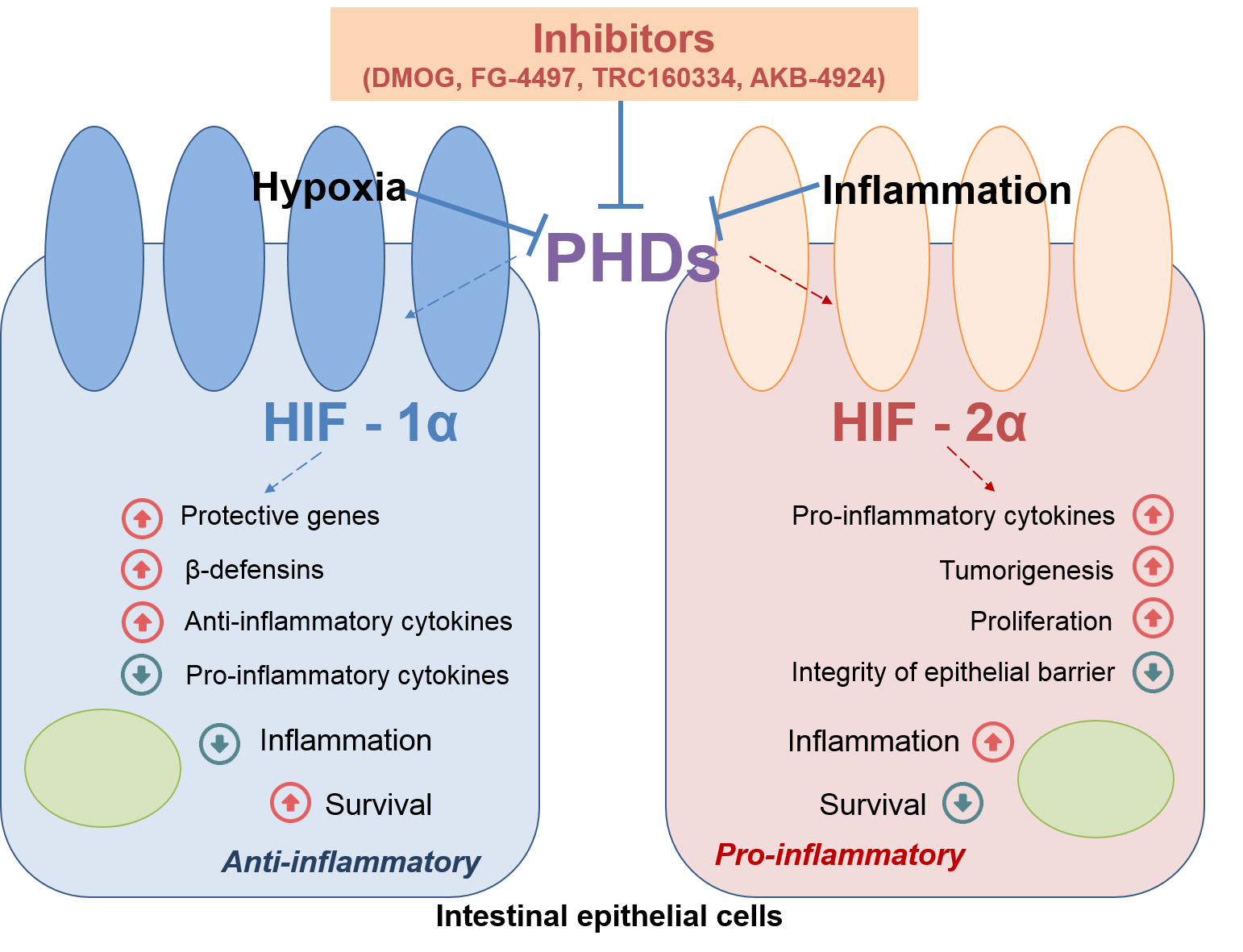
Figure 1. The role of HIFs in intestinal inflammation. PHDs – Prolyl Hydroxylase Domain Proteins, Using of PHDs inhibitors leads to non-selective, temporary activation of HIF-1α and HIF-2α.
According to the literature, in patients with CD and UC, expression of HIF-1α and HIF-2α is increased in colonocytes [32]. Moreover, a positive correlation was detected between the expression of HIF-1α and the severity of the disease course in patients with UC during remission and in the acute phase of diseases [34].
The most comprehensive study of the expression of HIF-1α and HIF-2α, as well as the effect on the organism of PHD inhibitors in connection with systemic disorders in humans, is difficult for several reasons; therefore, these studies are widely conducted in experimental models.
The so-called “chemical” models of IBD are most often used in experimental studies. Oral or rectal administration of specific chemical agents results in intestine inflammation that is similar in clinical and morphological manifestations to UC or CD. Such chemical agents are, for example, sodium dextran sulfate (Dextran Sodium Sulfate, DSS), TNBS (2,4,6-Trinitrobenzenesulfonic acid), oxazolone [35].
The role of hypoxia-inducible factors in the development of IBD and UC, as well as their influence on the severity of the inflammatory process, is primarily investigated in experimental models. HIF-1α knockout mice have been shown to be more susceptible to TNBS colitis [36], but knockout of this gene does not affect the severity of DSS colitis. The low expression of HIF-2α reduces the severity of the DSS colitis, despite the increase in the production of pro-inflammatory cytokines [32]. While overexpression of HIF-2α leads to a more severe course of UC, spontaneous inflammation of the intestine [32], as well as progression of cancer in experimental models [37]. An increase in the severity of DSS colitis was shown in mice with the deficiency of VHL that stabilizes HIF-1α and HIF-2α [38].
A lot of articles were devoted to the understanding the role of mechanisms of oxidative stress, in particular, PHD inhibitors, in search for the new drugs for the IBD treatment. Chemical inhibition of PHD, which both results in HIF-1α and HIF-2α subunits stabilization, does not affect the severity of UC, but instead leads to its remission through the HIF-1α-dependent pathway [31]. These data may seem contradictory; we discussed above, the activation of HIF-1α does not reduce the severity of the pro-inflammatory response induced by HIF-2α [32]. However it is possible to select a dose of dimethyloxalylglycine (DMOG), sufficient to activate HIF-1α, but not inducing inflammation along the HIF-2α-dependent pathway [32]. Also, usage of PHD inhibitors leads to non-selective, temporary activation of HIF-1α and HIF-2α, but not to prolonged overexpression of HIF-2α, that was demonstrated the induces of a spontaneous inflammatory process development in the intestine.
Thus, to sum up, HIF-1α is an anti-inflammatory agent in IBD and it ensures the integrity of the epithelial barrier and antimicrobial immunity by increasing the synthesis of β-defensins, while HIF-2α activates pro-inflammatory cytokine production and epithelial proliferation.
Perspectives for the treatment of ulcerative colitis
On the one hand, “physiological” hypoxia is a marker of the normal functioning of the intestine and, on the other hand, oxygen-dependent pathways and HIF play important role in the development of IBDs. Obviously, the search for new molecular targets for the pharmacological correction of oxygen-dependent signaling pathways attracts the attention of researchers in terms of developing new drugs for the treatment of IBDs.
Independent studies revealed the protective effect of various PHD inhibitors in many experimental models of colitis in 2008. To date, some pharmacological “hypoxia mimetics” have been developed, used to activate the HIF pathway for therapeutic effect. Many of these drugs today are non-specific hydroxylase inhibitors structurally related to 2-hydroxyglutarate [8, 38]. PHD inhibitors such as DMOG, FG-4497 and TRC160334, AKB-4924 were tested as therapeutic agents in colitis experimental models [38]. DMOG and FG-4497 are panhydroxylase inhibitors capable of inhibiting the activity of HIF-PHD and asparaginyl hydroxylase.
The authors first identified the protective effects of hydroxylase inhibition in models of colitis, in which the HIF-signaling pathways and the regulating inflammation nuclear factor NF-κB, probably plays an important role [39-42]. These two factors are interconnected, thus, on the one hand, in the proximal part of the HIF-1α gene promoter is the NF-κB-binding site [43] and, on the other hand, mRNA and HIF-1α protein levels increase dependent on NF-κB activation (the data obtained on the system inflammatory response induced by lipopolysaccharide) [44].
However therapeutic agents mentioned above are not specific and the result of their action is not only in HIF-1α subunit stabilization, but also in HIF-2α, which can enhance the inflammatory response. Recently it was investigated that, unlike HIF-1α, in the structure of the HIF-2α subunit there is a ligand-binding cavity. Despite the fact that endogenous substrates have not yet been identified, this difference in the structure of subunits is considered as a promising line of research for highly specific low molecular weight inhibitors as drugs for the control of IBDs in general and in particular UC [45].
Conclusion
Thus, UC is a disease, characterized by the interraption of the intestinal barrier, a disorder of immunity and microbial dysbiosis. Hypoxia is a key feature of the normal physiology of intestine, and in inflammatory conditions, such as UC, tissue hypoxia may be aggravated. HIF plays a central role in the normal function of a healthy intestinal mucosa, and using of PHD inhibitors are protective in experimental models of colitis. However, they are not specific and can enhance the inflammatory response in UC. In this case, it is important to continue to develop additional therapeutic strategies that use the protective potential of oxygen-sensitive methods, taking into account differences in spatial organization of HIF-1α and HIF-2α subunits.
Conflict of interest
The authors declare that they have no conflict of interest.
- Lukyanova LD. Cellular mechanism responsible for beneficial effects of hypoxic therapy. In: Adaptation biology and Medicine, Volume 3. J. Мoravec, et al., eds. New Dehli: Narosa Puplishing House, 2002: 290-303.
- Lyzhko NA. Molecular-genetic mechanisms of initiation, promotion and progression of tumors. Russian Journal of Biotherapy 2017; 16(4): 7-17. Russian. https://doi.org/10.17650/1726-9784-2017-16-4-7-17.
- Cinel I, Opal CM. Molecular biology of inflammation and sepsis: a primer. Crit Care Med 2009; 37(1): 291-304. https://doi.org/10.1097/CCM.0b013e31819267fb.
- Belousova EA. Ulcerative colitis and Crohn’ disease. Moscow: Triada, 2002; 128 p. Russian.
- Kapuller LL. Pathological changes of the colon in non-specific inflammatory bowel disease. In: Nonspecific inflammatory bowel disease. G.I. Vorob’ev, I.L. Khalif, eds. Moscow, Russia: Miklosh, 2008: 71-105. Russian.
- Dzhalilova DS, Polyakova MA, Diatroptov ME, Zolotova NA, Makarova OV. Morphological changes in the colon and composition of peripheral blood lymphocytes in acute colitis in mice with different resistance to hypoxia. Molecular Medicine 2018; 16(6): 46-50. Russian. https://doi.org/10.29296/24999490-2018-06-08.
- Kosyreva AM, Dzhalilova DS, Tsvetkov IS, Diatroptov ME, Makarova OV. Age-specific features of hypoxia tolerance and intensity of lipopolysaccharide-induced systemic inflammatory response in Wistar rats. Bull Exp Biol Med 2019; 166(5): 699-703. https://doi.org/10.1007/s10517-019-04421-3.
- Cummins EP, Crean D. Hypoxia and inflammatory bowel disease. Microbes Infect 2017; 19(3): 210-221. https://doi.org/10.1016/j.micinf.2016.09.004.
- Cioffi M, Rosa AD, Serao R, Picone I, Vietri MT. Laboratory markers in ulcerative colitis: Current insights and future advances. World J Gastrointest Pathophysiol 2015; 6(1): 13-22. https://doi.org/10.4291/wjgp.v6.i1.13.
- Ordas I, Eckmann L, Talamini M, Baumgart DC, Sandborn WJ. Ulcerative colitis. Lancet 2012; 380(9853): 1606-1619. https://doi.org/10.1016/S0140-6736(12)60150-0.
- Maeda S, Hsu LC, Liu H, Bankston LA, Iimura M, Kagnoff M. et al. Nod2 mutation in Crohn's disease potentiates NF-kappaB activity and IL-1beta processing. Science 2005; 307(5710): 734-738. https://doi.org/10.1126/science.1103685.
- Du X, Shi H, Li J, Dong Y, Liang J, Ye J, et al. Mst1/Mst2 regulate development and function of regulatory T cells through modulation of Foxo1/Foxo3 stability in autoimmune disease. J Immunol 2014; 192(4): 1525-1535. https://doi.org/10.4049/jimmunol.1301060.
- Neefjes J, Jongsma ML, Paul P, Bakke O. Towards a systems understanding of MHC class I and MHC class II antigen presentation. Nat Rev Immunol 2011; 11(12): 823-836. https://doi.org/10.1038/nri3084.
- Chyornenkaya TV. Pseudomembranous colitis: diagnosis, treatment and prevention. Russian Sklifosovsky Journal "Emergency Medical Care" 2016; (1): 33-39. Russian. https://www.elibrary.ru/item.asp?id=25778788.
- Trivedi PP, Jena GB. Ulcerative colitis-induced hepatic damage in mice: studies on inflammation, fibrosis, oxidative DNA damage and GST-P expression. Chem Biol Interact 2012; 201(3): 19-30. https://doi.org/10.1016/j.cbi.2012.12.004.
- Keith B, Johnson RS, Simon MC. HIF1alpha and HIF2alpha: sibling rivalry in hypoxic tumour growth and progression. Nat Rev Cancer 2011; 12(1): 9-22. https://doi.org/10.1038/nrc3183.
- Schödel J, Mole DR, Ratcliffe PJ. Pan-genomic binding of hypoxia-inducible transcription factors. Biol Chem 2013; 394(4): 507-517. https://doi.org/10.1515/hsz-2012-0351.
- Ivan M, Kondo K, Yang H, Kim W, Valiando J, Ohh M, et al. HIFalpha targeted for VHL-mediated destruction by proline hydroxylation: implications for O2 sensing. Science 2001; 292(5516): 464-468. https://doi.org/10.1126/science.1059817.
- Jaakkola P, Mole DR, Tian YM, Wilson MI, Gielbert J, Gaskell SJ, et al. Targeting of HIF-alpha to the von Hippel-Lindau ubiquitylation complex by O2-regulated prolyl hydroxylation. Science 2001; 292(5516): 468-472. https://doi.org/10.1126/science.1059796.
- Maxwell PH, Wiesener MS, Chang GW, Clifford SC, Vaux EC, Cockman ME, et al. The tumour suppressor protein VHL targets hypoxia-inducible factors for oxygen-dependent proteolysis. Nature 1999; 399(6733): 271-275. https://doi.org/10.1038/20459.
- Bohlen HG. Intestinal tissue pO2 and microvascular responses during glucose exposure. Am J Physiol 1980; 238(2): H164-H171. https://doi.org/10.1152/ajpheart.1980.238.2.H164.
- Matheson PJ, Wilson MA, Garrison RN. Regulation of intestinal blood flow. J Surg Res 2000; 93(1): 182-196. https://doi.org/10.1006/jsre.2000.5862.
- Hallbäck DA, Hultén L, Jodal M, Lindhagen J, Lundgren O. Evidence for the existence of a countercurrent exchanger in the small intestine in man. Gastroenterology 1978; 74(4): 683-690. https://www.ncbi.nlm.nih.gov/pubmed/631505.
- Merga Y, Campbell BJ, Rhodes JM. Mucosal barrier, bacteria and inflammatory bowel disease: possibilities for therapy. Dig Dis 2014; 32(4): 475-483. https://doi.org/10.1159/000358156.
- Ross MH, Pawlina W. Histology: A text and atlas: with correlated cell and molecular biology. Baltimore, MD: Lippincott Wiliams & Wilkins, 2006; 906 p.
- O'Hara AM, Shanahan F. The gut flora as a forgotten organ. EMBO Rep 2006;7(7): 688-693. https://doi.org/10.1038/sj.embor.7400731.
- Baumgart DC, Sandborn WJ. Crohn's disease. Lancet 2012; 380(9853): 1590-1605. https://doi.org/10.1016/S0140-6736(12)60026-9.
- Hsiao EY, McBride SW, Hsien S, Sharon G, Hyde ER, McCue T, et al. Microbiota modulate behavioral and physiological abnormalities associated with neurodevelopmental disorders. Cell 2013; 155(7): 1451-1463. https://doi.org/10.1016/j.cell.2013.11.024.
- Kelly CJ, Zheng L, Campbell EL, Saeedi B, Scholz CC, Bayless AJ, et al. Crosstalk between microbiota-derived short-chain fatty acids and intestinal epithelial HIF augments tissue barrier function. Cell Host Microbe 2015; 17(5): 662-671. https://doi.org/10.1016/j.chom.2015.03.005.
- Rivera-Chávez F, Zhang LF, Faber F, Lopez CA, Byndloss MX, Olsan EE, et al. Depletion of butyrate-producing Clostridia from the gut microbiota drives an aerobic luminal expansion of Salmonella. Cell Host Microbe 2016; 19(4): 443-454. https://doi.org/10.1016/j.chom.2016.03.004.
- Keely S, Campbell EL, Baird AW, Hansbro PM, Shalwitz RA, Kotsakis A, et al. Contribution of epithelial innate immunity to systemic protection afforded by prolyl hydroxylase inhibition in murine colitis. Mucosal Immunol 2014; 7(1): 114-123. https://doi.org/10.1038/mi.2013.29.
- Xue X, Ramakrishnan S, Anderson E, Taylor M, Zimmermann EM, Spence JR, et al. Endothelial PAS domain protein 1 activates the inflammatory response in the intestinal epithelium to promote colitis in mice. Gastroenterology 2013; 145(4): 831-841. https://doi.org/10.1053/j.gastro.2013.07.010.
- Xie L, Xue X, Taylor M, Ramakrishnan SK, Nagaoka K, Hao C, et al. Hypoxia-inducible factor/MAZ-dependent induction of caveolin-1 regulates colon permeability through suppression of occludin, leading to hypoxia-induced inflammation. Mol Cell Biol 2014; 34(16): 3013-3023. https://doi.org/10.1128/MCB.00324-14.
- Xu C, Dong W. Role of hypoxia-inducible factor-1α in pathogenesis and disease evaluation of ulcerative colitis. Exp Ther Med 2016; 11(4): 1330-1334. https://doi.org/10.3892/etm.2016.3030.
- Alex P, Zachos NC, Nguyen T, Gonzales L, Chen TE, Conklin LS, et al. Distinct cytokine patterns identified from multiplex profiles of murine DSS and TNBS-induced colitis. Inflamm Bowel Dis 2009; 15(3): 341-352. https://doi.org/10.1002/ibd.20753.
- Karhausen J, Furuta GT, Tomaszewski JE, Johnson RS, Colgan SP, Haase VH. Epithelial hypoxia-inducible factor-1 is protective in murine experimental colitis. J Clin Invest 2004; 114(8): 1098-1106. https://doi.org/10.1172/JCI21086.
- Xue X, Taylor M, Anderson E, Hao C, Qu A, Greenson JK, et al. Hypoxia-inducible factor-2alpha activation promotes colorectal cancer progression by dysregulating iron homeostasis. Cancer Res 2012; 72(9): 2285–2293. https://doi.org/10.1158/0008-5472.CAN-11-3836.
- Shah YM. The role of hypoxia in intestinal inflammation. Mol Cell Pediatr 2016; 3(1): 1. https://doi.org/10.1186/s40348-016-0030-1.
- Taylor CT, Colgan SP. Regulation of immunity and inflammation by hypoxia in immunological niches. Nat Rev Immunol 2017; 17(12): 774-785. https://doi.org/10.1038/nri.2017.103.
- Fratantonio D, Cimino F, Speciale A, Virgili F. Need (more than) two to Tango: Multiple tools to adapt to changes in oxygen availability. Biofactors 2018; 44(3): 207-218. https://doi.org/10.1002/biof.1419.
- Krzywinska E, Stockmann C. Hypoxia, metabolism and immune cell function. Biomedicines 2018; 6(2). pii: E56. https://doi.org/10.3390/biomedicines6020056.
- Stothers CL, Luan L, Fensterheim BA, Bohannon JK. Hypoxia-inducible factor-1α regulation of myeloid cells. J Mol Med (Berl) 2018; 96(12): 1293-1306. https://doi.org/10.1007/s00109-018-1710-1.
- van Uden P, Kenneth NS, Rocha S. Regulation of hypoxia-inducible factor-1alpha by NF-kappaB. Biochem J 2008; 412(3): 477-484. https://doi.org/10.1042/BJ20080476.
- Frede S, Stockmann C, Freitag P, Fandrey J. Bacterial lipopolysaccharide induces HIF-1 activation in human monocytes via p44/42 MAPK and NF-kappaB. Biochem J 2006; 396(3): 517-527. https://doi.org/10.1042/BJ20051839.
- Scheuermann TH, Li Q, Ma HW, Key J, Zhang L, Chen R, et al. Allosteric inhibition of hypoxia inducible factor-2 with small molecules. Nat Chem Biol 2013; 9(4): 271-276. https://doi.org/10.1038/nchembio.1185.
Received 04 September 2019, Accepted 13 January 2020
© 2019, Postovalova E.A., Makarova O.V., Kosyreva A.M., Dzhalilova D.S.
© 2019, Russian Open Medical Journal
Correspondence to Anna M. Kosyreva. Address: Tsyurupa str., 3, Moscow, 117418, Russia. Phone: +79269501760. Email: kosyreva.a@list.ru.